Grasa de sobrepaso y eCG en sincronización de ovejas / Pérez Ruiz et al. (2025)
Inclusión de grasa de sobrepaso y uso de eCG sobre las variables reproductivas en un protocolo de sincronización en ovejas de pelo subnutridas
Inclusion of fat by pass and use of eCG on reproductive variables in a synchronization protocol in undernourished hair sheep
Elizabeth Pérez Ruiz1, Miguel Paul Conde Hinojosa2, Jaime Gallegos Sánchez3, Glafiro Torres Hernández3,
Yolanda Osorio Marín4*
*Autor para correspondencia: osorio.marin.yolanda@hotmail.com Fecha de recepción: 8 de octubre de 2024
Fecha de aceptación: 26 de enero de 2025 Fecha de publicación: 31 de enero de 2025
1Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, C.E. La Campana, Chihuahua, México.
2Instituto de Ciencias Agropecuarias, Universidad Autónoma del Estado de Hidalgo, Tulancingo de Bravo, México.
3Colegio de Postgraduados Campus Montecillo, Montecillo, Texcoco, Estado de México, México.
4Unidad Académica Metztitlán, Universidad Politécnica de Francisco I. Madero, Metztitlán, Hidalgo, México.
![]()
ISSN electrónico: 2992-7501 2(2), 27 - 39 (Ene - Jun 2025)
https://doi.org/10.29059/cvpa.v2i2.27
Resumen
El objetivo de este estudio fue evaluar el efecto de la suplementación con grasa de sobrepaso y el uso de eCG en un protocolo de sincronización en ovejas de pelo subnutridas lactantes y vacías. Un total de 112 ovejas de pelo: 55 lactantes y 57 vacías con un peso promedio de 38±8 kg fueron asignadas a uno de cuatro tratamientos. Se utilizó un diseño completamente al azar; donde se evaluó la suplementación con grasa de sobrepaso (con y sin) y el uso de 300 UI de eCG (con y sin). Las variables evaluadas fueron: respuesta al estro, tiempo de inicio al estro, tasa de retorno, tasa de gestación, tasa de parto, prolificidad y fecundidad. La suplementación con grasa de sobrepaso y la administración de eCG no modificaron la respuesta al estro (p < 0.05 ), tasa de gestación, prolificidad o fecundidad en ovejas desnutridas lactantes y no lactantes. Sin embargo, se comprobó que las ovejas lactando responden al protocolo de sincronización iniciado al día 25 postparto, esta respuesta fue similar a la obtenida en el grupo de ovejas vacías.
Palabras clave: anestro postparto, desnutrición, nutrición focalizada.
Abstract
The objective of this study was to evaluate the effect of bypass fat supplementation and the use of eCG in a synchronization protocol in undernourished lactating and empty hair ewes. A total of 112 hair sheep: 55 lactating and 57 empty with 38±8 kg mean weight were assigned to one of the four treatments. A completely randomized design was used, where was evaluated the addition of bypass fat (with and without) and 300 IU of eCG (with and without). Ewes in all treatment responded adequately to hormonal treatment for estrus synchronization. Bypass fat supplementation and eCG administration did not modify estrus response, pregnancy rate, prolificacy or fecundity in lactating and non-lactating undernourished ewes. However, it was found that lactating ewes respond to the synchronization protocol initiated on day 25 postpartum.
Keywords: postpartum anestrous, focus nutrition, un dernutrition.
Introducción
La producción de ovinos en México se realiza en todo el país, principalmente en la zona centro. De acuerdo con datos oficiales, el inventario nacional ovino es de 8.8 millones; y poco más de 40 % de esta población ovina se encuentra en la región centro del país, en el Estado de México, Hidalgo, Veracruz y Puebla (SIAP, 2023). Además, la demanda de carne de ovino también se concentra en estos estados, aproximadamente 85 % de la carne de ovino que se consume a nivel nacional (Díaz-Sánchez, 2018).
En México, las unidades de producción ovina (UPO) están ubicadas en distintas zonas geográficas. El desarrollo de estas unidades productivas está condicionada por distintos factores tales como disponibilidad de recursos, mercado, condiciones socioeconómicas, acceso a la tierra, disponibilidad de insumos y tecnología. Predominan las de tipo extensivo y familiar, este tipo de unidad de producción también es denominado como tradicional (Hernández-Bautista et al., 2022). La alimentación en estas UPO se basan en el pastoreo a orilla de caminos vecinales, terrenos comunales y pastizales en zonas de conservación (Bobadilla-Soto et al., 2021). La producción ovina a pequeña escala, de tipo tradicional, tiene un papel relevante para las familias involucradas en esta actividad, puesto que les permite aprovechar los subproductos y residuos de las cosechas (Herrera-Haro et al., 2019). Entre las desventajas del sistema de producción extensivo en la ovinocultura mexicana, se pueden mencionar la fluctuación estacional en la disponibilidad de ganado para el abasto, irregularidad en el tipo y la condición de los animales que se producen (Vázquez-Martínez et al., 2009); así como bajos índices productivos y reproductivos (Herrera-Haro et al., 2019). Adicionalmente, otra desventaja en los sistemas extensivos, es la disponibilidad de alimento, sobre todo en las etapas importantes para los eventos reproductivos, como la gestación y la lactancia (Silva et al., 2022).
Los ovinos en pastoreo están expuestos a periodos de subnutrición, debido a los cambios en la composición nutricional de los pastizales en las diferentes estaciones del año. Estos cambios en la disponibilidad de alimento tienen una influencia negativa en diferentes procesos relacionados con el eje reproductivo. El déficit nutricional puede incrementar la duración del anestro postparto o estacional, así como comprometer el establecimiento de la gestación (Meikle et al., 2018). La restricción de nutrientes durante la gestación lleva a un balance energético negativo (BEN) en la oveja, lo cual impactará en varios procesos relacionados con la secreción láctea, crecimiento del feto, desarrollo de la cría postparto y finalmente el reinicio de la actividad reproductiva postparto (Rekik et al., 2010). Si esta escasez coincide con el último tercio de gestación de las ovejas, es inevitable la movilización de reservas corporales de la madre para asegurar el desarrollo y crecimiento fetal, garantizando la permanencia del producto hasta el momento del parto, sin embargo, los estados de desnutrición de la madre pueden causar alteraciones en el desempeño de productivo y la salud de la descendencia (Sinclair et al., 2016).
Uno de los métodos para mejorar el comportamiento reproductivo de las ovejas es la suplementación estratégica o nutrición focalizada (Martin, 2022). En ovinos, es posible estimular el desarrollo folicular al incrementar el consumo de energía (Scaramuzzi et al., 2011). Al mejorar estratégicamente la calidad de la dieta y las prácticas de alimentación, es posible incrementar la tasa de ovulación, mejorar la implantación embrionaria y, por lo tanto, la eficiencia reproductiva (Juengel et al., 2021; Martin, 2022; Tesfaye et al., 2023).
Estas estrategias cobran especial interés para solventar las necesidades de los animales en los sistemas de producción extensiva, cuando los requerimientos nutricionales no pueden ser cubiertos sólo con el pastoreo. Ejemplos de la estrategia de nutrición focalizada con ingredientes energéticos, es el uso de la grasa de sobrepaso (Bianchi et al., 2018). Además, incluir otras estrategias de manejo, como el destete precoz o amamantamiento controlado, ha mostrado su utilidad en la reducción del anestro postparto (Morales-Terán et al., 2004; Ungerfeld & Sanchez-Davila, 2012).
Adicionalmente, el uso de hormonales, como la gonadotropina coriónica equina (eCG, por sus siglas en inglés: equine corionic gonadotropin) puede mejorar la fecundidad (Dias et al., 2020; Kridli & Al-Khetib, 2006). En rumiantes, las eCG se une a los receptores de FSH y LH, estimulando el crecimiento folicular; y durante la época no reproductiva (anestro), la aplicación de esta hormona mejora la respuesta de estro y la ovulación (Kridli & Al-Khetib, 2006). En la época reproductiva, la dosis de eGC recomendada es de 200 a 300 UI, y se observa una mejor sincronización del estro y un menor tiempo de inicio al estro (después del retiro del progestágeno) con dosis de 300 UI (Quintero et al., 2022).
El objetivo de este estudio fue evaluar la respuesta a un protocolo de sincronización de estros, con la inclusión de grasa de sobrepaso, como estrategia de nutrición focalizada y la aplicación de 300 UI eCG en ovejas lactantes y vacías subnutridas.
MATERIALES Y MÉTODOS
Ubicación del área de estudio
El estudio se realizó en el Laboratorio de Reproducción de Ovinos y Caprinos (LaROCa) del Campus Montecillo, Colegio de Postgraduados, Texcoco, Estado de México, ubicado a una latitud N de 19º 27´51´´ y 98º 54´34´´de longitud O. La región se encuentra a 2250 msnm y presenta clima templado con una precipitación media anual de 644.8 mm. La temperatura promedio es de 15ºC (García, 2004). El protocolo de sincronización inició durante la época reproductiva (octubre) y los partos se registraron en abril.
Animales, manejo y tratamientos
Para este estudio se respetaron los lineamientos del Reglamento para el uso y cuidado de animales destinados a la investigación en el Colegio de Postgraduados (2016), de acuerdo con las normas oficiales Mexicanas NOM-024-ZOO-1995, NOM-033- ZOO-1995 y NOM-051-ZOO-1995. Se utilizaron 112 ovejas de pelo como animales experimentales. Estas fueron divididas en dos grupos. El Grupo 1, conformado por 55 ovejas lactantes (con 25 días postparto); Grupo 2, conformado por 57 ovejas vacías (no lactantes). El peso vivo promedio de los animales experimentales fue de 38 ± 8 kg y una condición corporal de 2.2 ± 0.04 para las ovejas lactantes y 1.8 ± 0.03, para las ovejas vacías (no lactantes). El peso vivo de los animales experimentales de este estudio estaba por debajo del promedio del rebaño (40.7 ± 7.1 kg), así como CC debajo de 2.5, en una escala de 1 a 5 (González-Maldonado et al., 2023), por lo cual estos animales fueron clasificados como subnutridos. Las ovejas de los dos grupos fueron reasignadas a uno de los cuatro tratamientos por grupo, estableciendo un total de 8 grupos experimentales. Se utilizó un diseño completamente al azar, con la adición de grasa de sobrepaso (Grasa y Sin grasa) y la aplicación de eCG (eCG y Sin eCG). Para el caso de las ovejas vacías, los tratamientos fueron: T1: V+eCG (Ovejas vacías + 300 UI de eCG; n= 14), T2: V+G (Ovejas vacías + 8% de grasa de sobrepaso en dieta; n=14), T3: V+G+eCG (Ovejas vacías + 8% de grasa de sobrepaso incluida en la dieta + 300 UI de eCG; n=14) y T4: V (Ovejas vacías, sin grasa y sin eCG; n=15). Para las ovejas lactantes los tratamientos fueron similares a los empleados en las ovejas vacías: T1: L+eCG (Ovejas lactantes + 300 UI de eCG; n= 15), T2: L+G (Ovejas lactantes + 8% de grasa de sobrepaso en dieta; n=13), T3: L+G+eCG (Ovejas lactantes + 8% de grasa de sobrepaso incluida en la dieta + 300 UI de eCG; n=12) y T4: L (Ovejas lactantes, sin grasa ni eCG; n=15).
Alimentación
Todas las ovejas fueron alimentadas según sus requerimientos, con una dieta balanceada y de acuerdo con su estado fisiológico; lactantes (2.3 Mcal kg-1 de MS de energía metabolizable y 18% de proteína cruda) y vacías (1.9 Mcal kg-1 de MS de energía metabolizable y 15% de proteína cruda) de acuerdo con lo recomendado por el NRC (2007) (Tabla 1). Para los tratamientos que incluían la grasa de sobrepaso, se agregó el 8% de grasa granulada PALMALIFE® a la dieta base (misma que recibieron las ovejas sin grasa de sobrepaso). El agua se proporcionó ad líbitum durante todo el estudio.
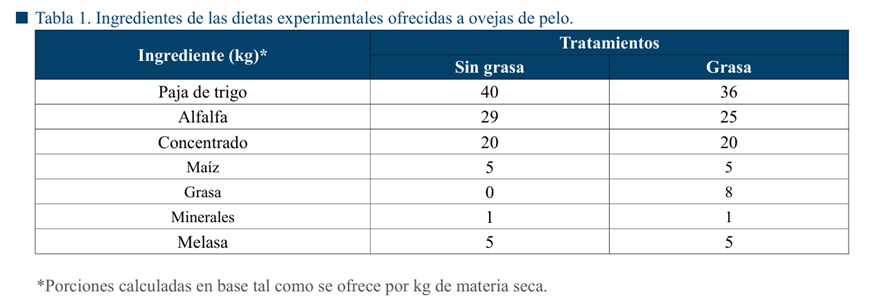
El protocolo de sincronización duró 10 días y la dieta experimental con grasa de sobrepaso (estrategia de nutrición focalizada) se ofreció durante 5 días. Entre los días 5 y 9 del protocolo de sincronización para favorecer el desarrollo folicular. En el caso de las ovejas que se encontraban lactando, los corderos permanecieron con las ovejas en amamantamiento continuo. Para complementar la alimentación de los corderos, nacidos durante la primera semana de octubre, a partir del día 7 de nacimiento, se les proporcionó un concentrado peletizado (creep feeding) a libre acceso.
Protocolo de sincronización del estro
El protocolo de sincronización en los dos grupos de ovejas (vacías y lactantes) se realizó de manera simultánea. En las ovejas lactantes, el dispositivo intravaginal (CIDR, Controlled Internal Drug Release; con 0.3 g de progesterona, Pfizer) fue colocado el día 25 postparto. El tratamiento con progesterona se realizó por 9 días. 48 h antes del retiro del CIDR se aplicó 1 mL oveja-1 de prostaglandina (PGF2α, Dinoprost; Lutalyse, Pharmacia & Upjhon, Michigan, USA), por vía intramuscular, con la finalidad de lisar cualquier cuerpo lúteo presente. Las ovejas en los tratamientos con eCG, recibieron 300 UI (Folligon, Intervet, D.F., México), por vía intramuscular 48 h antes del retiro del CIDR. La detección de estros se realizó después de retirar el CIDR, con la ayuda de machos celadores cada 4 h, por un periodo de 72 h.
Inseminación artificial (IA)
Antes de la inseminación, las ovejas fueron rasuradas y se desinfectó la región abdominal, especialmente en la base de la ubre, donde se realizaron las incisiones. El semen utilizado para la inseminación se colectó de carneros sanos y aptos reproductivamente. Las ovejas que mostraron signos de estro fueron inseminadas por laparoscopia, 12 o 18 horas posteriores al inicio del estro. Se utilizaron pajillas de 0.25 mL, con una concentración espermática de 50 x10-6, en cada cuerno uterino se colocó la mitad de la pajilla de semen.
Diagnóstico de gestación
El diagnóstico de gestación se realizó por vía transrectal al día 35 post-inseminación con un ultrasonido portátil (Aloka SSD 500, transductor 7 Mhz).
Las variables evaluadas fueron las siguientes:
Respuesta al estro. Número de ovejas que manifestaron signos de estro después del retiro del dis positivo respecto al número total de ovejas de cada tratamiento.
![]()
Inicio al estro. Intervalo de
tiempo entre el retiro del CIDR y el inicio del estro. La distribución se
determinó con el número de ovejas que entraron en estro hasta las 72 horas
posteriores al retiro del CIDR.
Retorno al estro. Número de ovejas que manifestaron signos de estro en los dos ciclos posteriores a la IA (17 o 34 días post-inseminación).
![]()
Tasa de gestación. Número de ovejas diagnosticadas gestantes, con respecto al total de ovejas por tratamiento.
![]()
Tasa de parto. Número de ovejas paridas respecto al número total de ovejas por tratamiento.
![]()
Prolificidad. Número de corderos nacidos vivos o muertos con respecto al número de ovejas pari das por tratamiento.
![]()
Fecundidad. Número total de corderos nacidos vivos o muertos con respecto al número total de ovejas por tratamiento.
![]()
Análisis estadístico
Para el análisis estadístico se utilizó el paquete estadístico SAS 9.0 (2012). Para las variables respuesta al estro, retorno al estro, tasa de gestación y tasa de parto se utilizó regresión logística, mediante el procedimiento PROC LOGISTIC. Las variables prolificidad y fecundidad, se analizaron mediante una distribución POISSON, con el PROC GENMOD. Para la variable inicio al estro se realizó la prueba de Shapiro & Wilk, para observar normalidad univariada y posteriormente se realizó el análisis con el método de tiempos de vida, usando el estimador de Meier (Kaplan-Meier), con el procedimiento LIFETEST. Las diferencias se determinaron con la prueba de Log-Rank. Para todas las variables se consideraron diferencias significativas cuando el valor fue (p < 0.05).
RESULTADOS
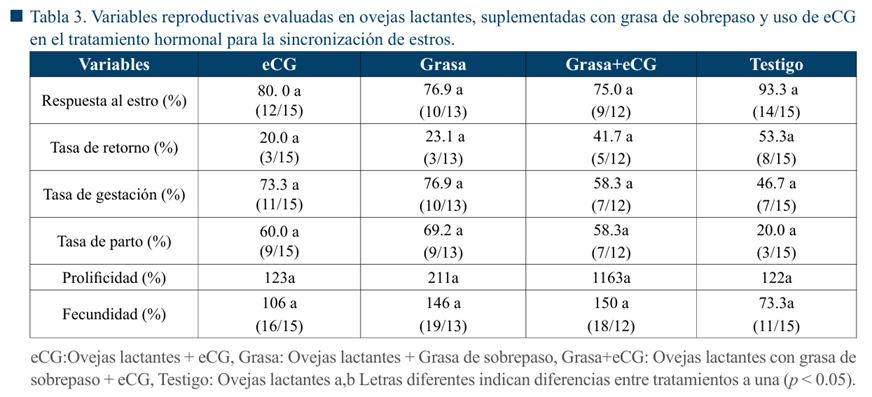
En las Tablas 2 y 3 se muestran los resultados obtenidos, de acuerdo con el estado fisiológico de las ovejas. En ninguna de las variables evaluadas se observaron diferencias significativas (p > 0.05) debidas a la inclusión de grasa de sobrepaso en la dieta o la aplicación de eCG.

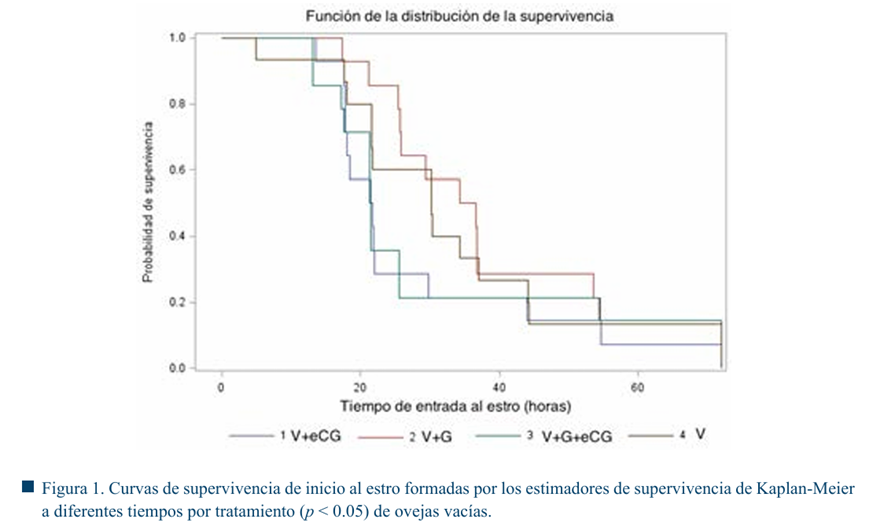
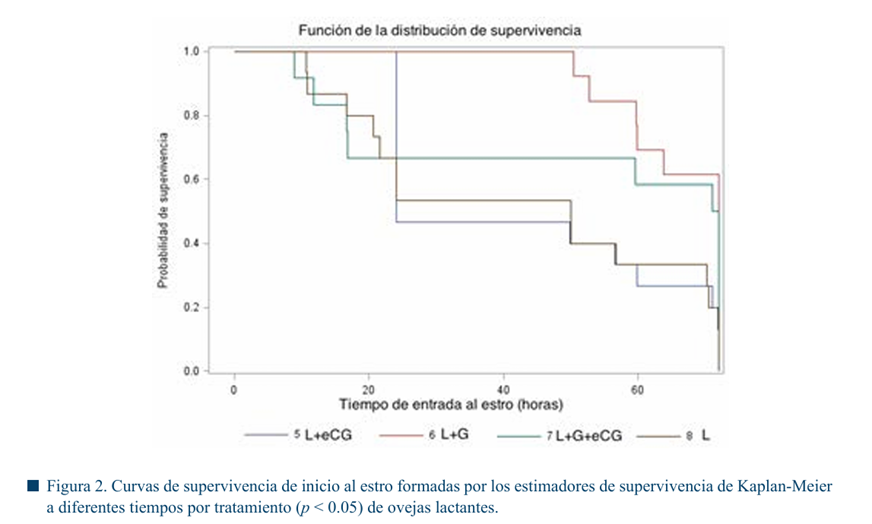
En las Figuras 1 y 2 se muestran los tiempos de inicio al estro, no se observaron diferencias significativas entre tratamientos (p > 0.05). Sin embargo, en el grupo de ovejas vacías el inicio del estro se agrupó en un lapso de tiempo menor (32.5 h), en comparación con las ovejas lactantes (50.3 h) en promedio.
DISCUSIÓN
Se sabe que el estado nutricional es un aspecto clave para el desempeño reproductivo de las ovejas, y los requerimientos aumentan en momentos como el último tercio de gestación y la lactación (Dos Santos et al., 2017). En diversos estudios se ha reportado que las ovejas con condición corporal (CC) alta tienen un mejor desempeño reproductivo, que puede ser medido en términos de fertilidad, actividad folicular y mayor tasa de ovulación, en comparación con ovejas en CC baja (Delgadillo & Martin, 2015; Viñoles et al., 2014). Entre los hallazgos que se han descrito en ovejas con CC alta vs CC baja, se encuentran diferencias en la duración del estro, a favor de aquellas con CC alta (De la Isla et al., 2010); presencia de folículos de mayor tamaño cuando la CC es alta (Viñoles et al., 2002) y menor tasa de ovulación asociada a una CC baja (De la Isla et al., 2010). En el caso de las ovejas lactantes, además existen otros factores que también influyen negativamente en el reinicio de la actividad reproductiva, entre ellos el efecto del amamantamiento, a través de la producción de péptidos opioides endógenos (POEs), neurotransmisores implicados en el mecanismo neuroendocrino que impide la ovulación durante el periodo postparto (Arroyo et al., 2009). Sin embargo, algunos estudios han mostrado que la CC alta o baja no modifica el reinicio de la actividad ovárica y ciclicidad en ovejas de pelo lactantes (Orozco-Durán et al., 2022). En este estudio, los animales experimentales se encontraban con una CC menor a 2.5 (escala 1 a 5) el porcentaje de respuesta al protocolo de sincronización de ovejas (vacías y lactantes) fue similar al reportado por otros autores (González- Maldonado et al., 2023) en ovejas con condiciones corporales más altas.
El uso de eCG es efectivo para estimular el desarrollo folicular en protocolos de sincronización de estros (Boscos et al., 2002); el 100% de las ovejas vacías a las que se les aplicó eCG mostraron signos de estro aún cuando su condición corporal fue menor a 2.5. Las ovejas lactantes del tratamiento con grasa de sobrepaso + eCG en ovejas lactantes mostraron el porcentaje de respuesta más bajo (75 %). Esto puede deberse al efecto de la lactancia, diversos estudios han evidenciado que la prolactina desempeña un papel como modulador del sistema reproductivo en las ovejas. Esto se debe a su capacidad para inhibir la liberación de LH y FSH en la hipófisis, así como la liberación de la hormona gonadotropina a nivel hipotalámico, esto influye en la regulación de las hormonas reproductivas (Morales-Terán et al., 2004; Yue et al., 2024). Una estrategia comúnmente recomendada para mejorar el desempeño reproductivo de las ovejas es la nutrición focalizada de corta duración. Al mejorar el consumo de nutrientes, es posible modificar la secreción de factores de crecimiento y el flujo de nutrientes, por lo que la viabilidad de los folículos dependientes de gonadotropinas puede mejorar (Scaramuzzi et al., 2011). En un estudio con ovejas Pelibuey desnutridas, la alimentación por 21 días con una ración del 120% de los requerimientos recomendados por el NRC, modificó la tasa de ovulación, tasa de retorno al estro, tasa de parición y prolificidad, los autores concluyen que en ovejas desnutridas son altamente sensibles a los aportes nutricionales positivos (González-Maldonado et al., 2023). Sin embargo, estos resultados difieren de los reportados por Viñoles et al. (2010), quienes mencionan que, aunque las ovejas con CC baja tienen folículos sensibles a gonadotropinas, estos no responden a los cambios metabólicos de la nutrición focalizada, por lo que no se observaron cambios en la tasa de ovulación en ovejas con CC baja, suplementadas con granos de lupino por 6 d-1. A pesar de que las ovejas en este estudio mostraron signos de estro en respuesta al protocolo de sincronización, la suplementación con grasa de sobre paso y/o la aplicación de eCG, no mejoraron ninguna de las variables evaluadas (Tablas 1 y 2). Existe la posibilidad de que el estado metabólico de estas ovejas influyera negativamente en la calidad de los folículos ováricos, la competencia de los ovocitos, así como el desarrollo embrionario, por lo que un porcentaje considerable de ovejas en todos los tratamientos retornaron al estro y no quedaron gestantes. También es posible que el periodo de suplementación y la ración ofrecida fueran insuficientes para provocar cambios en el estado metabólico de estas ovejas y, por tanto, no se observaron efectos positivos en las variables evaluadas. Por otra parte, entre las ventajas del uso de la eCG se pueden mencionar el efecto positivo en el crecimiento folicular, dependiendo de la dosis, puede mejorar la tasa de ovulación, tasa de concepción y favorecer la presentación de partos múltiples (Kridli & Al-Khetib, 2006). Además, al estimular el crecimiento folicular, la eCG, favorece la agrupación del estro (Boscos et al., 2002) y este se presenta en menor tiempo (Quintero-Elisea et al., 2011). Sin embargo, en este estudio no se observó ningún efecto de la aplicación de eCG, en el tiempo de inicio al estro en ovejas vacías o lactantes (Figura 1 y 2). Es posible que, debido plano de desnutrición de las ovejas experimentales, no se observaron efectos significativos de la eCG para la tasa de gestación, prolificidad y fecundidad (p > 0.05). La desnutrición puede disminuir o inhibir la secreción pulsátil de LH ya sea en hembras o en machos (Merkley et al., 2020) y reducir la liberación de GnRH hacia la eminencia media y en la circulación portal hipofisiaria (I’Anson et al., 2000). La Kispeptina es un neuropéptido esencial para reproducción (Meikle et al., 2018) y en ovejas jóvenes, la suspensión de alimento por tres días resulta en una reducción moderada en la secreción de LH y Kispeptina. En ovejas adultas ovariectomizadas, un estado de desnutrición crónica, puede causar la supresión en la secreción de LH y la reducción del ARNm de kispeptina a nivel núcleo arcuato (Backholer et al., 2010). Es posible que, en este estudio, la producción y secreción de kispeptina se vio afectada, por la suplementación con grasa o aplicación de eCG. Además, la restricción en el consumo de alimento también puede producir un mal funcionamiento del rumen y disminución en la síntesis de proteína microbiana, que contribuye a mantener un balance negativo (Cal-Pereyra et al., 2012). Estos estados de desnutrición tienen consecuencias a largo plazo en la descendencia, las deficiencias de energía pueden afectar la productividad de las crías, en cuanto a crecimiento, producción de lana, leche o carne (Sinclair et al., 2016). Y pueden resultar críticos para los corderos, debido a que la tasa de mortalidad puede ser muy elevada durante los primeros días de nacidos (Banchero et al., 2004). En los últimos años, los conceptos de “Programación fetal” y “Origen de la salud y enfermedad en el desarrollo” (DoHaD, por su acrónimo en inglés), han cobrado una gran importancia tanto en lo relacionado a la salud humana (Hsu & Tain, 2021) como de los animales de producción (Sinclair et al., 2016). Los estados de desnutrición en la oveja pueden afectar la productividad futura de su descendencia, por lo que la producción de lana, carne y leche, así como el desempeño reproductivo, se pueden ver comprometidos (Martin, 2022). En los sistemas de producción basados en el pastoreo extensivo, como es el caso de la producción ovina en México, la nutrición es uno de los principales desafíos, puesto que la disponibilidad de forraje de buena calidad está sujeta a variaciones estacionales. Por lo anterior, las estrategias de nutrición focalizada en estos sistemas de producción, con ingredientes de bajo costo, pueden ser una alternativa para mejorar el desempeño de estos animales en momentos clave del desarrollo. Los resultados obtenidos en este estudio muestran que la suplementación con grasa de sobre paso no mejoró la tasa de ovulación, prolificidad o la fecundidad; sin embargo, se recomienda continuar con esta línea de investigación, para evaluar otros aspectos de la desnutrición en las características reproductivas de ovejas en condiciones de subnutrición, así como las implicaciones en el desempeño de los corderos.
CONCLUSIONES
La suplementación con grasa de sobrepaso y la administración de eCG no modificaron la respuesta al estro, tasa de gestación, prolificidad o fecundidad en ovejas desnutridas lactantes y no lactantes. Sin embargo, se comprobó que las ovejas lactando responden de manera favorable al protocolo de sincronización iniciado al día 25 postparto, por lo que es factible el uso de un protocolo de sincronización antes de los 60 días postparto.
REFERENCIAS
Arroyo, J., Magaña-Sevilla, H., & Camacho-Escobar, M. A. (2009). Regulación neuroendocrina del anestro posparto en la oveja. Tropical and Subtropical Agroecosystems, 10(3), 301-312.
Backholer, K., Smith, J. T., Rao, A., Pereira, A., Iqbal, J., Ogawa, S., Li, Q., & Clarke, I. J. (2010). Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide Y and proopiomelanocortin cells. Endocrinology, 151(5), 2233-2243. https://doi.org/10.1210/en.2009-1190
Banchero, G. E., Quintans, G., Martin, G. B., Lindsay, D. R., & Milton, J. T. B. (2004). Nutrition and colostrum production in sheep. 1. Metabolic and hormonal responses to a high-energy supplement in the final stages of pregnancy. Reproduction, Fertility and Development, 16(6), 633. https://doi.org/10.1071/RD03091
Bianchi, A. E., Macedo, V. P., Da Silva, A. S., da Silveira, A. L. F., Hill, J. A. G., Zortéa, T., Rossi, R. M., & Batista, R. (2018). Effect of the addition of protected fat from palm oil to the diet of dairy sheep. Revista Brasileira de Zootecnia, 47, e20160137. https://doi.org/10.1590/rbz4720160137
Bobadilla-Soto E.E., Ochoa-Ambriz F., & Perea-Peña M. (2021). Dinámica de la producción y consumo de carne ovina en México 1970 a 2019. Agronomía Mesoamericana, 32(3), 963-982. https://doi.org/10.15517/am.v32i3.44473
Boscos, C. M., Samartzi, F. C., Dellis, S., Rogge, A., Stefanakis, A., & Krambovitis, E. (2002). Use of progestagen–gonadotrophin treatments in estrus synchronization of sheep. Theriogenology, 58(7), 1261-1272. https://doi.org/10.1016/S0093-691X(02)01040-3
Cal-Pereyra, L., Acosta-Dibarrat, J., Benech, A., Da-Silva, S., Martín, A., & González-Montańa, J. R. (2012). Toxemia de la gestación en ovejas. Revisión. Revista Mexicana de Ciencias Pecuarias, 3(2), 247-264.
Colegio de Postgraduados. (2022). Reglamento para el uso y cuidado de animales destinados a la investigación. https://www.colpos.mx/wb_pdf/norma_interna/Reglamento_COBIAN-CP_2023.pdf
De la Isla, G., Aké-López, R., Ayala-Burgos, A., & González-Bulnes, A. (2010). Efecto de la condición corporal y la época del año sobre el ciclo estral, estro, desarrollo folicular y tasa ovulatoria en ovejas Pelibuey mantenidas en condiciones de trópico. Veterinaria México, 41(3), 167-175.
Delgadillo, J. A., & Martin, G. B. (2015). Alternative methods for control of reproduction in small ruminants – a focus on the needs of grazing animal industries. Animal Frontiers, 5(1), 57-65. https://doi.org/10.2527/af.2015-0009
Dias, J. H., Miranda, V. O., Oliveira, F. C., Vargas Junior, S. F., Haas, C. S., Costa, V. G.G., Lucia Jr., T., Vieira, A. D., Corcini, C. D., & Gasperin, B. G. (2020). Treatment with eCG and hCG to induce onset of estrous cycles in ewes during the non-breeding season: Efects on follicular development and fertility. Animal Reproduction Science, 212, 106-232. https://doi.org/10.1016/j.anireprosci.2019.106232
Díaz-Sánchez, C. C., Jaramillo-Villanueva, J. L., Bustamante-González, Á., Vargas-López, S., Delgado-Álvarado, A., Hernández-Mendo, O., &. Casiano-Ventura, M. Á. (2018). Evaluation of the profitability and competitiveness of sheep production systems in the region of Libres, Puebla. Revista Mexicana de Ciencias Pecuarias, 9(2), 263-277. https://doi.org/10.22319/rmcp.v9i2.4495
Dos Santos, S., Falbo, M., Sandini, I., Ishiy L., Deliberalli, & Posso, L. (2017). Ryegrass pasture supplementation strategies for lactating ewe performance. Turkish Journal Veterinary Animal Science, 41(6), 787-792. https://doi.org/10.3906/vet-1703-86
García, E. (2004). Modificaciones al sistema de clasificación Climática de Köppen (5ta ed.). Instituto de Geografía-UNAM.
González-Maldonado, J., Tejeda Sartorius, O., Ramírez-Ramírez, A. I., & Gallegos-Sánchez, J. (2023). Reproductive performance of hair sheep under different body conditions and feeding levels. Pesquisa Agropecuária Brasileira, 58, e03092. https://doi.org/10.1590/S1678-3921. pab2023.v58.03092
Hernández-Bautista, J., Rodríguez-Magadán, H. M., Salinas, R. T., Aquino, C. M., & Mariscal M. A. (2022). Caracterización de los sistemas de producción familiar ovina en la Mixteca Oaxaqueña, México. Revista Mexicana Ciencias Pecuaria, 13(4), 1009-1024. https://doi.org/10.22319/rmcp.v13i4.6100
Herrera-Haro, J. G., Álvarez-Fuentes, G., Bárcena-Gama, R., & Núñez-Aramburu, J. M. (2019). Caracterización de los rebaños ovinos en el sur del Distrito Federal, México. Acta Universitaria, 29, e2022. doi. http://doi.org/10.15174.au.2019.2022
Hsu, C. N., & Tain, Y. L. (2021). Animal models for DOHaD research: Focus on hypertension of developmental origins. Biomedicines, 9(6), 623. https://doi.org/10.3390/biomedicines9060623
I’Anson, H., Manning, J. M., Herbosa, C. G., Pelt, J., Friedman, C. R., Wood, R. I., Bucholtz, D. C., & Foster, D. L. (2000). Central inhibition of gonadotropin-releasing hormone secretion in the growth-restricted hypogonadotropic female sheep. Endocrinology, 141(2), 520-527. https://doi.org/10.1210/endo.141.2.7308
Juengel, J. L., Cushman, R. A., Dupont, J., Fabre, S., Lea, R. G., Martin, G. B., Mossa, F., Pitman, J. L., Price, C. A., & Smith, P. (2021). The ovarian follicle of ruminants: the path from conceptus to adult. Reproduction Fertility and Development, 33(10), 621-642. https://doi.org/10.1071/RD21086
Kridli, R. T. & Al-Khetib, S. S. (2006). Reproductive responses in ewes treated with eCG or increasing doses of royal jelly, Animal Reproduction Science, 92(1-2), 75-85. https://doi.org/10.1016/j.anireprosci.2005.05.021
Martin, G. B. (2022). Frontiers in sheep reproduction – making use of natural responses to environmental challenges to manage productivity. Animal Reproduction, 19(4), e20220088. https://doi.org/10.1590/1984-3143AR2022-0088
Meikle, A., Brun, V. de, Carriquiry, M., Soca, P., Sosa, C., Adrien, M. de L., Chilibroste, P., & Abecia, J. A. (2018). Influences of nutrition and metabolism on reproduction of the female ruminant. Animal Reproduction, 15(Suppl. 1), 899-911. https://doi.org/10.21451/1984-3143-AR2018-0017
Merkley, C. M., Shuping, S. L., & Nestor, C. C. (2020). Neuronal networks that regulate gonadotropin-releasing hormone/luteinizing hormone secretion during undernutrition: Evidence from sheep. Domestic Animal Endocrinology, 73, 106469. https://doi.org/10.1016/j.domaniend.2020.106469
Morales-Terán, G., Pro-Martínez, A., Figueroa-Sandoval, B., Sánchez-del-Real, C., & Gallegos-Sánchez, J. (2004). Amamantamiento continuo o restringido y su relación con la duración del anestro postparto en ovejas Pelibuey. Agrociencia, 38, 165-171.
NRC. (2007). Nutrient requirements of small ruminants: Sheep, goats, cervids, and New World camelids. National Academies Press. https://doi.org/10.17226/11654
Orozco Durán, K. E, Segura Correa, J., Herrera Ojeda, J., Herrera Camacho, J., & Bottini Luzardo, M. B. (2022). Body condition score at lambing did not affect the resumption of ovarian and estrous activity in lactating Pelibuey ewes. Nova Scientia, 14(28), 1-9. https://doi.org/10.21640/ns.v14i28.2965
Quintero, E. J., Macías, C. U., Alvarez, V. D., Correa, C. A., González, R. A., Lucero, M. F., Soto, N. S., & Avendaño, R. L. (2011). The effects of time and dose of pregnant mare serum gonadotropin (PMSG) on reproductive efficiency in hair sheep ewes. Tropical Animal Health and Production, 43, 1567-1573. https://doi.org/10.1007/s11250-011-9843-z
Quintero, E J., Olguín, A. H., Velázquez, M. J., Garay,M. J., Vázquez, R, L., Limas, M. A., & Cancino, S. J. (2022). Effect of breed, breeding season, eCG dose, and eCG application time on the estrous cycle of hair ewe lambs. Agro Productividad, 15(7), 187-194. https://doi.org/10.32854/agrop.v15i7.2321
Rekik, M., Ben Salem, H., Lassoued, N., Chalouati, H., & Ben Salem, I. (2010). Supplementation of Barbarine ewes with spineless cactus (Opuntia ficus-indica f. inermis) cladodes during late gestation-early suckling: Effects on mammary secretions, blood metabolites, lamb growth and postpartum ovarian activity. Small Ruminant Research, 90(1-3), 53-57. https://doi.org/10.1016/j.smallrumres.2009.12.051
SAS. (2012). SAS/STAT: User´s guide statistics released 9.1. (2nd ed.). SAS Institute Inc.
Scaramuzzi, R. J., Baird, D. T., Campbell, B. K., Driancourt, M. A., Dupont, J., Fortune, J. E., Gilchrist, R. B., Martin, G. B., McNatty, K. P., McNeilly, A. S., Monget, P., Monniaux, D., Viñoles, C., & Webb, R. (2011). Regulation of folliculogenesis and the determination of ovulation rate in ruminants. Reproduction Fertility and Development, 23(3), 444-467. https://doi.org/10.1071/RD09161
Servicio de Información Agroalimentaria y Pesquera (SIAP). (2023). Población ganadera. https://nube.siap.gob.mx/poblacion_ganadera/
Silva, S. R., Sacarrão-Birrento, L., Almeida, M., Ribeiro, D. M., Guedes, C., González Montaña, J. R., Pereira, A. F., Zaralis, K., Geraldo, A., Tzamaloukas, O., Cabrera, M. G., Castro, N., Argüello, A., Hernández-Castellano, L. E., Alonso-Diez, Á. J., Martín, M. J., Cal-Pereyra, L. G., Stilwell, G., & de Almeida, A. M. (2022). Extensive sheep and goat production: The role of novel technologies towards sustainability and animal welfare. Animals, 12(7), 885. https://doi.org/10.3390/ani12070885
Sinclair, K. D., Rutherford, K. M. D., Wallace, J. M., Brameld, J. M., Stöger, R., Alberio, R., Sweetman, D. S., Gardner, D., Perry, V. E. A., Adam, C. L., Ashworth, C. J., Robinson, J. E., Dwyer, C. M. (2016). Epigenetics and developmental programming of welfare and production traits in farm animals. Reproduction, Fertility and Development, 28(10), 1443-78. https://doi.org/10.1071/RD16102
Tesfaye, A., Asmare, B., Abiso, T., & Wamatu, J. (2023). Effect of nutritional flushing using long-term energy and protein supplementation on growth performance and reproductive parameters of Doyogena ewes in Ethiopia. Veterinary Science, 10(6), 368. https://doi.org/10.3390/vetsci10060368
Ungerfeld, R., & Sanchez-Davila, F. (2012). Oestrus synchronization in postpartum autumn-lambing ewes: Effect of postpartum time, parity, and early weaning. Spanish Journal of Agricultural Research, 10(1), 62. https://doi.org/10.5424/sjar/2012101-233-11
Vázquez-Martínez, I., Vargas, L. S., Zaragoza, R. J. L., Bustamante, G. A., Calderón, S. F., Rojas, A. J., & Casiano, V. M. A. (2009). Tipología de explotaciones ovinas en la sierra norte del estado de Puebla. Técnica Pecuaria México, 47(4), 357-369.
Viñoles, C., Paganoni, B. L., McNatty, K. P., Heath, D. A., Thompson, A. N., Glover, K. M. M., Milton, J. T. B., & Martin, G. B. (2014). Follicle development, endocrine profiles and ovulation rate in adult Merino ewes: effects of early nutrition pre- and post-natal; and supplementation with lupin grain. Reproduction, 147(1), 101-110. https://doi.org/10.1530/REP-13-0104
Viñoles, C., Forsberg, M., Banchero G., Rubianes E. (2002). Ovarian follicular dynamics and endocrine profiles in Polwarth ewes with high and low body condition. Animal Science, 74(3), 539-545. https://doi.org/10.1017/S1357729800052693
Viñoles, C., Paganoni, B., Glover, K. M. M, Milton, J. T. B., Blache, D., Blackberry, M. A., & Martin, G. B. (2010). The use of a ‘first-wave’ model to study the effect of nutrition on ovarian follicular dynamics and ovulation rate in the sheep. Reproduction, 140(6), 865-874. https://doi.org/10.1530/REP-10-0196
Yue, S., Chen, J., Duan, C., Li, X., Yang, R., Chen, M., Li, Y., Song, Z., Zhang, Y., & Liu, Y. (2024). The effect of prolactin on gene expression and the secretion of reproductive hormones in ewes during the estrus cycle. Animals, 14(13), 1873. https://doi.org/10.3390/ani14131873