Estrategias de nutrición y alimentación para reducir las emisiones de metano en rumiantes bajo sistemas pastoriles: Una revisión
Nutrition and feeding strategies to reduce methane emissions in ruminants under pastoral systems: A review
Luz Yosahandy Peña-Avelino1, Jorge Alva-Perez1, Gerardo Noé Rosales-Matínez1,
Said Hernández-Contreras1, Ivonne Ceballos-Olvera1*
Autor para correspondencia:iceballos@docentes.uat.edu.mx Fecha de recepción: 11 de enero de 2024
Fecha de aceptación: 25 de enero de 2024 Fecha de publicación: 31 de enero de 2024
1Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas. Ciudad Victoria, Tamaulipas, México.
![]()
ISSN electrónico: 2992-7501 1(2), 60 - 73 (Ene - Jun 2024)
https://doi.org/10.29059/cvpa.v2i2.18
Resumen
Los sistemas de producción de rumiantes en pastoreo mantienen un potencial de mitigación de gases de efecto invernadero (GEI), en especial el metano entérico (CH4). En sistemas intensivos de producción de leche bovina se estima una emisión de 1 kg de CO2eq kg de energía-leche corregida (ECM), comparada con > 7 kg de CO2eq kg de ECM en los sistemas extensivos. El objetivo de este artículo fue evaluar las estrategias usados para mitigar las emisiones de CH4 y destacar aquellas con potencial de implementación en los sistemas pastoriles, desde el punto de vista económico y sustentable. Las estrategias de alimentación y modificadores de la biota en el rumen pueden disminuir las emisiones en un rango de 14 a 50%, mientras que reducciones por manejo y selección de rumiantes van de 15 a 30%. La adopción de más de una estrategia en los sistemas pastoriles puede ayudar a mitigar las emisiones de CH4.
Palabras clave: metano, impacto ambiental, producción animal, agostaderos.
Abstract
Grazing ruminant production systems maintain a potential for greenhouse gas (GEI) mitigation, especially enteric methane (CH4). In intensive bovine milk production systems, an emission of 1 kg CO2eq kg energy-corrected milk (ECM) is estimated, compared to > 7 kg CO2eq kg ECM in extensive systems. The objective of this article was to evaluate the strategies used to mitigate CH4 emissions and to highlight those with potential for implementation in pastoral systems, from an economic and sustainable point of view. Feeding strategies and rumen biota modifiers can reduce emissions by 14 to 50%, while reductions from ruminant management and selection range from 15 to 30%. Adopting more than one strategy in pastoral systems can help mitigate CH4 emissions.
Keywords: methane, environmental impact, animal production, pastures.
Introducción
El sector ganadero mundial contribuye con 7.1 gigatoneladas (Gt) de dióxido de carbono equivalente (CO2-eq por año), que constituyen 14.5% de las emisiones inducidas por los humanos (Gerber et al., 2013). Los principales gases de efecto invernadero (GEI) emitidos a la atmósfera son el óxido nitroso (N2O), metano (CH4) y dióxido de carbono (CO2). El N2O y el CH4 son gases de efecto invernadero que retienen calor proveniente del sol y presentan un potencial de calentamiento global de 265 y 28 veces mayor que el CO2, en un tiempo medio de 100 años (IPCC, 2013). La presencia de estos gases corresponde a pérdidas de nitrógeno (N), energía y materia orgánica que afectan la eficiencia y productividad en los sistemas pecuarios. El CH4 es producido por la fermentación entérica de los rumiantes y la descomposición de la materia orgánica del estiércol. El 87% de CH4 se produce en el rumen, mientras el 13% restante en el colon (Murray et al., 1976), este se expulsa a través de gases y eructos. El N2O proviene del N presente en el estiércol y los fertilizantes químicos. Se estima que para 2050 la demanda de carne y leche aumentará en 73% y 58%, en relación con el 2010 (Garg, 2012). Esto representará un impulso en el sector ganadero e incrementará la competencia por los recursos naturales con otros sectores. Así, es necesario incorporar prácticas y tecnologías que aumenten la productividad y eficiencia del uso de los recursos naturales, al mismo tiempo en que mitigan la emisión de GEI. Hay tareas que se están realizando para reducir los daños al ambiente, sin embargo, no son suficientes. Los climas regionales se están transformando y esto dificultará la producción agropecuaria debido a la intensidad de sequias o inundaciones (Rojo-Martínez et al., 2003).
Los Análisis de Ciclo de Vida (ACV) han mostrado que la carne y leche producida a partir de cereales tiene un impacto ambiental menor que aquella que es producida en los sistemas que utilizan el forraje como alimento (Crosson et al., 2011; O´Mara, 2011). Esto se ha observado en Europa, América del Norte y Rusia con similares producciones de carne y leche, comparado con África y América Latina, pero con diferencia en la emisión de CH4 entérico de 26% contra 69% respectivamente (Hristov et al., 2013). En Europa, cerca del 80% de la carne de vacuno proviene de terneros y vacas de reemplazo de razas con alta producción de leche, lo que produce poca intensidad de emisiones. Por lo anterior, el objetivo de esta revisión fue analizar las estrategias de mitigación de CH4 entérico de rumiantes en sistemas intensivos que pudieran aplicarse a sistemas de producción pastoriles.
Metano en sistemas ganaderos de pastoreo
Se ha determinado que la cadena de suministro ganadero emite 2.0 Gt de CO2, equivalente (CO2 -eq), 3.1 Gt de CO2-eq de CH4 y 2.0 Gt de CO2-eq de N2O por año. La producción de carne y leche de vacuno contribuye con el 60% de las emisiones totales del sector pecuario, mientras que las emisiones en pequeños rumiantes ascienden al 6.5% (Gerber et al., 2013). El porcentaje de emisiones de GEI durante la producción de bovinos en las zonas templadas, tropicales y aridas es de 44%, 27% y 29%, respectivamente (Opio et al., 2013). Esto se justifica por una mayor concentración de hatos en las zonas templadas, debido a que las condiciones de producción son favorables para los bovinos especializados en dicha región.
En Asia, África, América Latina y el Caribe mantienen los niveles más altos de GEI (casi 1.3 Gt de CO2-eq) debido a la gran cantidad de ganado con productividad baja. Por lo tanto, estas regiones deben implementar acciones para disminuir la emisión de los GEI. Para lograr este objetivo se ha sugerido seguir diferentes estrategias como: alimentación; manejo de los pastizales; estructura del hato y genética (Gerber et al., 2013; Hristov et al., 2013). La implementación de un mejor manejo ayudará con la disminución en la emisión de los GEI, con énfasis en CH4 enterico. Esto se traducirá en un mejor rendimiento productivo y aprovechamiento de los recursos naturales.
Alimentación y modificadores de la biota ruminal
La adopción de prácticas que permitan maximizar la producción y utilización de los nutrientes contenidos en los alimentos ayuda a disminuir las emisiones de los GEI. La revisión de los atributos químicos en los alimentos permite conocer la calidad y formulación de una alimentación que satisfaga los requerimientos de los rumiantes a través de dietas mezcladas (Hristov et al., 2013). Esto es conocido como alimentación de precisión, la tasa de pasaje en el rumen se mejora al ser provisto de nutrientes en una cantidad ideal. Esto hace que los microoganismos del rumen incrementen su actividad y como consecuencia la digestión mejora (Macor et al., 2020).
Asi mismo, la dieta de los rumiantes tiene influencia en la emisión de GEI (Min et al., 2022); la inclusión de concentrados (granos o alimentos energéticos ≥ 40% en la dieta) modifica los parámetros ruminales. El pH del líquido ruminal se acidifica (≤ 5.8) y en consecuencia, la cantidad de bacterias aminolíticas se incrementa al promover la defaunación de protozoarios. La relación de acetato:propionato y la cantidad de CH4 disminuye (Posada-Ochoa et al., 2014). Sin embargo, debido a la competencia de los cereales con la alimentación humana y los incrementos por los combustibles fósiles. Esto ocasiona el incremento del costo del insumo y las emisiones de los GEI durante la cadena productiva. Por lo tanto, el consumo de pasturas de buena calidad no debe sustituirse por concentrado (Hristov et al., 2013).
Modificadores de la biota en el rumen
El uso de aditivos químicos y suplementos alimenticios en los rumiantes tiene efecto sobre la mitigación de CH4, porque se incrementa el ácido propiónico, las proteínas de sobrepaso, el balance energético, la degradación de los carbohidratos (almidón, sacarosa) y se controla la concentración de ácido láctico (Knapp et al., 2014). Mwenya et al. (2004) al evaluar diferentes cepas de levaduras (Trichosporum sericeum), bacterias lácticas (Leuconostoc mesenteroides) y de β1-4 galacto-olisacaridos encontraron potencial para disminuir las emisiones de CH4 en el rumen (Hristov et al., 2013).
La implementación de inhibidores halogenados tiene actividad antimetanogénica en la fermentación ruminal y en la estructura de las comunidades microbianas. En un estudio realizado por Abecia et al. (2012) con cabras lecheras, el bromoclorometano (BCM) produjo una disminución en la producción de gas CH4 en un 33% y un incremento en la producción de leche (36%). Sin embargo, el BCM destruye la capa de ozono, por eso se restringe el uso en países Europeos (Gerber et al., 2013; Hristov et al., 2013). Hristov et al. (2015) estudiaron el efecto de un inhibidor de la metil coenzima-M reductasa (CoM) llamado 3-nitrooxipropanol (3NOP) en vacas Holstein de alta productcción de leche (45 kg leche d-1), observaron que el uso de 3NOP (40-80 mg kg-1 MS) no afecta el consumo de materia seca, la producción de leche, la digestibilidad y además reduce las emisiones de CH4 por vaca al día hasta en 30%.
Los aceptores de electrones más estudiados, hasta el momento, son el fumarato, los nitratos, los sulfatos y el nitroetano que han reportado una disminución de CH4 hasta en un 50%. El ácido málico (AM) y fumárico (AF) pueden reducir la producción de CH4 (Gerber et al., 2013; Hristov et al., 2013). El efecto del AM dependerá de la dosis, la proporción forraje:concentrado y los métodos de evaluación, ya sea in vitro o in vivo (Foley et al., 2009). En tanto, AF tiene potencial para incrementar la glucogénesis y por tanto el rendimiento de leche, pero la cantidad debe ser regulada para evitar el riesgo de acidosis (Wood et al. 2009). Anderson et al. (2010) encontraron que el nitroetano en condiciones in vitro destina a la metanogénesis más de un 30% de los electrones. Brown et al. (2011) observaron una disminución del 26 al 24% de CH4 con dosis de nitroetano (60 y 120 mg kg-1 de peso vivo de novillos Holstein). El uso de nitratos puede ser una estrategia efectiva en los países en vías de desarrollo, porque la cantidad de nitratos por fertilización de pasturas se incrementaría y el N proveniente de los nitratos podría contribuir a mantener o incrementar la producción.
La adición de ionóforos a la dieta inhibe la producción de CH4, aumenta la permeabilidad de la membrana y el paso de los iones (Voet et al., 2007), porque modifican la población y el metabolismo de la flora intestinal, con una mejor digestión y absorción de los nutrientes (Nuñez, 2009). Kim et al. (2014) muestran que la monensina sódica (MSo) tiene efecto sobre la producción de CH4. En novillos productores de carne se observó un mayor efecto antimetanogenico, comparado con vacas productoras de leche. Sin embargo, se determinó que este efecto depende de la dosis del MSo, del alimento consumido y la composición de la dieta (Appuhamy et al., 2013). Diversos estudios muestran que la MSo presenta un efecto menos consistente en rumiantes alimentados con zacates (Hristov et al., 2013).
Los aceites de almendra, coco, girasol y linaza mezclados con dietas altas en forraje pueden ser útiles para reducir las emisiones de CH4 hasta en un 50% (Chuntrakort et al., 2014; Kobayashi, 2010). Estos aceites tienen efectos sobre la biota ruminal porque reducen la población de protozoarios, la cantidad producida de ácidos grasos volátiles de cadena corta y la digestibilidad de la fibra (Chung et al., 2011). Brask et al. (2013) observaron que la suplementación con aceite de colza entre 6 y 6.5% en vacas lecheras redujo la emisión de CH4 en un 14%, sin comprometer la digestibilidad y la producción de leche. Los aceites esenciales como tomillo, orégano, canela, ajo y rábano poseen bioactivos con actividad antibacteriana y cuentan con la capacidad de inhibir la metanogénesis hasta en un 72% (Benchaar & Greathead, 2011), sin embargo, son necesarias dosis altas y los estudios que se han realizado han sido hechas en condiciones in vitro (Jahani-Azizabadi et al., 2011).
Los ácidos grasos (AG) tienen el potencial para suprimir la metanogénesis ruminal. Existe una discusión entre los que presentan mayor eficacia: saturados o poliinsaturados. Patra (2013) observó una mayor efectividad con los AG poliinsaturados. No obstante, los resultados en diversos estudios muestran que grasas saturadas también pueden ser útiles para disminuir las emisiones de CH4. Grainger & Beauchchemin (2011) encontraron que un incremento de 10 g/kg-1 en la grasa dietaría provocó la disminución de gas CH4 en 1 g/kg-1 de alimento consumido en vacas lecheras y 2.6 g/kg-1 en pequeños rumiantes. Los modificadores de la biota ruminal son una estrategia viable en la disminución del CH4 enterico, sin embargo, es necesario realizar estudios sobre su efecto por más de un año y las dosis necesarias para ser implementados en sistemas pastoriles.
Sistemas de producción
Los sistemas de producción pastoriles tienen una amplia extension de terreno, donde los rumiantes obtienen su alimento de la vegetación silvestre o inducida (zacates). La productividad de los sistemas de pastoreo es insuficiente en términos de producción por animal y unidad de trabajo, pero alta en términos de producción a partir de recursos muy limitados como el agua y los cereales. El N2Ode evaluación, ya sea in vitro o in vivo (Foley et al., 2009). En tanto, AF tiene potencial para incrementar la glucogénesis y por tanto el rendimiento de leche, pero la cantidad debe ser regulada para evitar el riesgo de acidosis (Wood et al. 2009). Anderson et al. (2010) encontraron que el nitroetano en condiciones in vitro destina a la metanogénesis más de un 30% de los electrones. Brown et al. (2011) observaron una disminución del 26 al 24% de CH4 con dosis de nitroetano (60 y 120 mg kg-1 de peso vivo de novillos Holstein). El uso de nitratos puede ser una estrategia efectiva en los países en vías de desarrollo, porque la cantidad de nitratos por fertilización de pasturas se incrementaría y el N proveniente de los nitratos podría contribuir a mantener o incrementar la producción.
La adición de ionóforos a la dieta inhibe la producción de CH4, aumenta la permeabilidad de la membrana y el paso de los iones (Voet et al., 2007), porque modifican la población y el metabolismo de la flora intestinal, con una mejor digestión y absorción de los nutrientes (Nuñez, 2009). Kim et al. (2014) muestran que la monensina sódica (MSo) tiene efecto sobre la producción de CH4. En novillos productores de carne se observó un mayor efecto antimetanogenico, comparado con vacas productoras de leche. Sin embargo, se determinó que este efecto depende de la dosis del MSo, del alimento consumido y la composición de la dieta (Appuhamy et al., 2013). Diversos estudios muestran que la MSo presenta un efecto menos consistente en rumiantes alimentados con zacates (Hristov et al., 2013).
Los aceites de almendra, coco, girasol y linaza mezclados con dietas altas en forraje pueden ser útiles para reducir las emisiones de CH4 hasta en un 50% (Chuntrakort et al., 2014; Kobayashi, 2010). Estos aceites tienen efectos sobre la biota ruminal porque reducen la población de protozoarios, la cantidad producida de ácidos grasos volátiles de cadena corta y la digestibilidad de la fibra (Chung et al., 2011). Brask et al. (2013) observaron que la suplementación con aceite de colza entre 6 y 6.5% en vacas lecheras redujo la emisión de CH4 en un 14%, sin comprometer la digestibilidad y la producción de leche. Los aceites esenciales como tomillo, orégano, canela, ajo y rábano poseen bioactivos con actividad antibacteriana y cuentan con la capacidad de inhibir la metanogénesis hasta en un 72% (Benchaar & Greathead, 2011), sin embargo, son necesarias dosis altas y los estudios que se han realizado han sido hechas en condiciones in vitro (Jahani-Azizabadi et al., 2011).
Los ácidos grasos (AG) tienen el potencial para suprimir la metanogénesis ruminal. Existe una discusión entre los que presentan mayor eficacia: saturados o poliinsaturados. Patra (2013) observó una mayor efectividad con los AG poliinsaturados. No obstante, los resultados en diversos estudios muestran que grasas saturadas también pueden ser útiles para disminuir las emisiones de CH4. Grainger & Beauchchemin (2011) encontraron que un incremento de 10 g/kg-1 en la grasa dietaría provocó la disminución de gas CH4 en 1 g/kg-1 de alimento consumido en vacas lecheras y 2.6 g/kg-1 en pequeños rumiantes. Los modificadores de la biota ruminal son una estrategia viable en la disminución del CH4 enterico, sin embargo, es necesario realizar estudios sobre su efecto por más de un año y las dosis necesarias para ser implementados en sistemas pastoriles.
Sistemas de producción
Los sistemas de producción pastoriles tienen una amplia extension de terreno, donde los rumiantes obtienen su alimento de la vegetación silvestre o inducida (zacates). La productividad de los sistemas de pastoreo es insuficiente en términos de producción por animal y unidad de trabajo, pero alta en términos de producción a partir de recursos muy limitados como el agua y los cereales. El N2O proveniente del manejo de estiércol es reducido porque la mayor parte el estiércol se deposita en la pastura, además, contiene menor cantidad de proteína en heces. Los sistemas mixtos e intensivos están asociados con el uso de fertilizantes químicos y la proporción de concentrado en la dieta (Pinares-Patiño et al., 2009; Rivera & Chará, 2021).
Los sistemas intensivos de producción mantienen a los animales estabulados y la interacción del ganado con los pastizales es reducido. Su alimentación es de tipo corte y acarreo. Esto puede ocasionar alteraciones en el suelo, balance del flujo de nutrientes del sistema por la falta de retorno de nutrientes por la resiembra o depósito natural de estiércol. El manejo del estiércol ha ganado relevancia porque es una de las formas para disminuir el impacto ambiental en suelo, aire y agua, a través de lixiviados. Esto origina una reducción de emisiones de GEI durante el almacenamiento, procesamiento y aplicación (Steinfeld et al., 2006).
Un ejemplo del control, en el manejo del estiércol, son los países desarrollados como Canadá y EUA, estos cuentan con alta tecnificación (Pinos-Rodríguez et al., 2012). Según estimaciones, la cantidad de estiércol que se recicla para la generación de biogás es del 1% mundial (Thøy et al., 2009). Las emisiones provenientes de la gestión del estiércol son elevadas en América del Norte donde el 27% del estiércol del sector lechero se maneja en sistemas líquidos que producen cantidades mayores de emisiones de CH4. La producción, elaboración de piensos y el estiércol son fuente de emisión, junto con la fermentación entérica. En cambio, en las regiones de baja productividad, la fuente principal de emisiones es la fermentación entérica (Opio et al., 2013).
En los sistemas mixtos, los herbívoros pasan algunas horas del día en galerones resguardándose de la intemperie, ahí son alimentados con suplementos y alimentos concentrados. Otros sistemas son los silvopastoriles, con algunas variantes de tipo como: árboles dispersos en los potreros, cercas vivas, bancos forrajeros de leñosas, pasturas en callejones de especies leñosas, frutales o plantaciones forestales (Villanueva et al., 2010).
La actividad ganadera basada en sistemas silvopastoriles tiene el potencial de producir leche, carne y servicios ambientales como la reducción de CH4 (Króliczewska et al., 2023). En estos sistemas la gestión del agua, la genética y el manejo del hato contribuyen a la adaptación, el cambio y variabilidad climática. Un manejo apropiado del pastoreo y los movimientos del ganado contribuyen a incrementar la productividad del ecosistema, la biodiversidad y la mitigación del cambio climatico (Steinfeld & Gerber, 2010; Vigan et al., 2017). Los sistemas mixtos o silvopastoriles tienen alternativas para la mitigación de emisiones con el pago por servicios ambientales, la conservación de la fauna silvestre o el almacenamiento de carbono (Villanueva et al., 2010). La deposición del estiércol en los sistemas mixtos puede ser a través de lagunas de almacenamiento conectados a biodigestores o distribuidos a los pastizales.
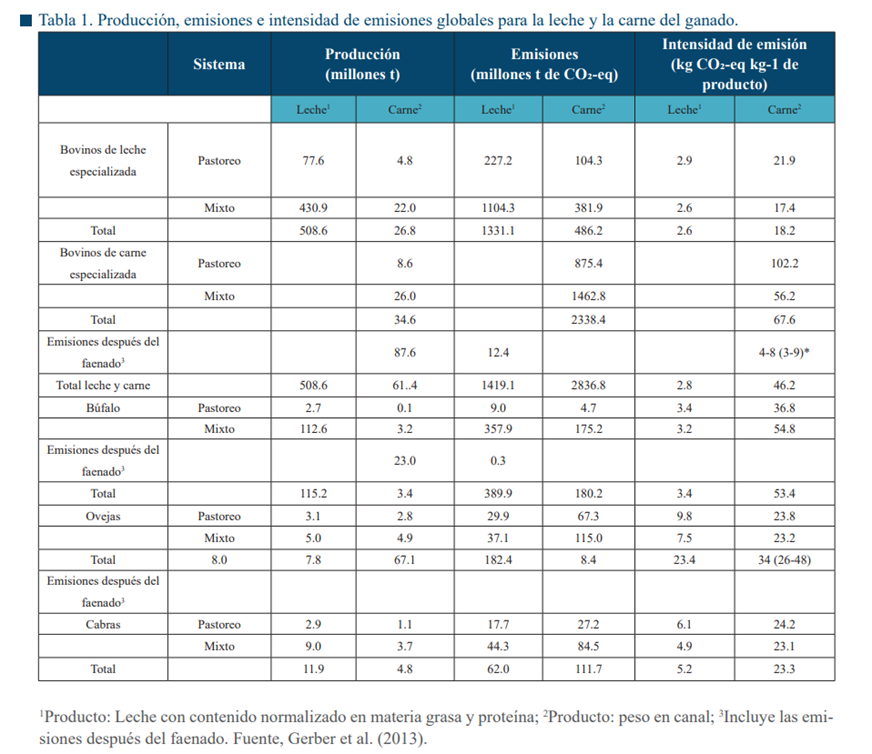
La intensidad de emisiones en los rumiantes es mayor en el sistema pastoril mientras que se reduce son un sistema mixto (Tabla 1). Si los sistemas pastoriles mantuvieran la misma producctividad que los sistemas mixtos, entonces habría un incremento en las emisiones de entre 12 y 20% por producción de leche y carne en los bovinos lecheros. La situación se agrava en los bovinos especializados en carne porque se observaría un incremento del 55% de las emisiones. En los sistemas mixtos, a pesar de generar una mayor cantidad de emisiones (GEI), tienen una mayor eficiencia reproductiva, manejo, faenado y la combinación de estos factores resulta en una menor intensidad de emisiones y mayor potencial de productividad (Opio et al., 2013).
En los sistemas pastoriles y silvopastoriles, la estimación de la capacidad de carga animal es un elemento necesario para el manejo ganadero sustentable (Holenchek et al., 1989). En nuestro país, la única referencia a esta estimación nacional fue la desarrollada por la Comisión Técnica Consultiva para la determinación de Coeficientes de Agostadero (COTECOCA, 1967), cuyos estudios, en su gran mayoría, datan de la época de los 60 y 70. La COTECOCA definió la capacidad de carga animal como el número de unidades animales (una unidad es una vaca gestante y lactante de 450 kg con su cría menor de 7 meses; en ovinos y caprinos: cinco hembras con su cría) que pueden pastorear en un potrero en un tiempo determinado, sin causar daño a la vegetación. Al respecto, la COTECOCA refiere que la condición de un agostadero o pastizal es excelente (76 y 100%) o buena (51 y 75%), con relación al porcentaje de cobertura vegetal. La aplicación de nuevas tecnologías mediante la utilización de drones permite estimar la capacidad de carga animal (Villa-Herrera et al., 2014).
El sobrepastoreo es un factor que incrementa las emisiones de CH4 debido a la disminución en la disponibilidad de forraje, esto fue observado por Baudracco et al. (2010) en vacas lecheras. Es conocido que la calidad de los forrajes y el aprovechamiento eficiente de los nutrientes en las dietas es una práctica que disminuye las emisiones de los GEI (Carmona-Flores et al., 2020). La dieta puede tener un impacto significativo en la química del estiércol y, por lo tanto, en las emisiones de los GEI. Dietas poco digestibles (< 40% fibra) generan emisiones elevadas de CH4 por unidad de energía ingerida, mientras que forrajes con alto contenido de azúcares podrían reducir la excreción de N urinario, la volatilización del amoníaco y las emisiones de N2O (Hristov et al., 2013). Las emisiones de CH4 pueden disminuir cuando se sustituye ensilado de pastos por ensilado de maíz (DEFRA, 2010; Van Gastelen et al., 2015).
En estudios realizados por Galindo et al. (2008) suplementaron vacas con 25% de Leucaena leucocephala como parte del consumo total de la materia seca y observaron incrementos en las poblaciones de bacterias celulolíticas, así como en la actividad específica de sus enzimas, mientras se redujo la población de protozoos ruminales. Montenegro & Abarca (2000) observaron que el pasto kikuyo (Pennissetum clandestinum) presentó una mayor eficiencia en la producción de leche comparado con el pasto estrella africana (Cynodon nlenfuensis), debido a la menor producción de CH4 que fue observada a los 28 días. En este sentido, la evaluación de los forrajes ha sido importante, Herrera-Pérez et al. (2023) encontraron que la producción acumulada de CH4 fue mayor en pasto estrella y pangola (Digita-riaeriantha Steud.) a los 30 días, mientras que a los 60 días las especies que mostraron mayor producción fueron Insurgente (Brachiaria brizantha Hochst. Stapf.), Mombaza (Panicummaximun Jacq. cv. Mombaza) y Pará (Brachiariamutica Stapf).
Por otro lado, estudios realizados en regiones cálidas, la asociación leguminosa y gramínea puede favorecer la disminución de los GEI debido a que las leguminosas tienen un mayor contenido de proteína, fibra y metabolitos secundarios como los polifenoles (taninos) que aumentan la digestibilidad de la dieta (Hristov et al., 2013). Plantas ricas en taninos y saponinas tienen el potencial para disminuir las emisiones de CH4. Los taninos (hidrolizables y condensados) han mostrado en diversos estudios in vitro e in situ actividad anti-metanógenica, aunque puede verse comprometido el consumo de alimento y el rendimiento productivo, es una estrategia recomendada (Goel & Makkar, 2012; Hess et al., 2003; Ku-Vera et al., 2020). Sin embargo, la efectividad para disminuir las emisiones de CH4 depende de la composición de las plantas (Jouany & Morgavi, 2007; Moscoso et al., 2017; Pellikaan et al., 2011).
Tiemann et al. (2008) explican que las leguminosas tropicales como Calliandra calothyrsus y Flemingia macrophylla, presentan contenidos de taninos que disminuyen la digestibilidad de la fibra y por tanto, se reduce la metanogénesis in vitro. Ramírez-Restrepo & Barry (2005) observaron que las emisiones de CH4 disminuyen con Chicorium intybus, Hedysarum coronarium y Lotus corniculatus, este último mostró una disminución del 21% asociado a un incremento en el consumo de alimento y producción de leche. Waghorn et al. (2002) evaluaron la producción de CH4 con diferentes forrajes y observaron que las emisiones disminuyeron un 16% en borregos que pastorearon sobre una mezcla de Lolium perenne y Trifolium repens, al adicionar pellets de Lotus pedunculatus (11.5 g CH4 kg-1 DM). Las saponinas tienen un efecto anti-protozoario debido al enlace de las saponinas con el colesterol de la membrana celular del protozoario, porque causa lisis e inhibe la metanogénesis (Díaz Puentes, 2009). Hess et al. (2003) menciona que el uso de frutos de Sapindus saponaria disminuyó las emisiones de CH4 (11%) en dietas con pastos de baja calidad con o sin suplementación de leguminosa. Abreu et al. (2003), señalaron que el uso del mismo árbol en proporciones de 8% de fruto no mostró efectos sobre la disminución de las emisiones de CH4. La producción de CH4 in vitro con niveles de 1.2 a 3.2 g de saponina L se redujo en un 44%, según lo indicado por Lila et al. (2003).
Galindo et al. (2005) encontraron efectos defaunantes con Gliricidia sepium, Sapindus saponaria, Arachis pintoi, S. aterrimum y Leucaena leucocephala, además de la existencia de una relación inversa entre el número de protozoos y el número de zoosporas móviles de hongos en el rumen. También, una relación inversa entre los contenidos de taninos condensados y de polifenoles totales con la degradabilidad de la materia seca. El efecto de S. saponaria mejoró el metabolismo energético por concepto de producción de metano. Galindo et al. (2011) indicaron que la suplementación con 20% en base seca de Tithonia diversifolia en la dieta disminuyó la población de protozoos en el rumen 2.7 veces y con ello la producción de CH4. Delgado et al. (2007) al analizar Morus alba y Trichanthera gigantea con respecto al forraje de Pennisetum observaron una mejora en el valor nutricional de la dieta y una disminución de CH4 (27%). Mao et al. (2010), en su experimento con borregos alimentados con 60% de pasto (Aneurolepidium Chinese Kitagawa) y 40% concentrado (base seca), observaron que cuando se adicionó al alimento saponinas provenientes de la hoja del té verde (Camellia sinensis) hubo una reducción. El uso de árboles y arbustos forrajeros es una alternativa económica y viable que permite incorporar al sistema una mayor carga animal manteniendo diferentes especies, esto ayuda a la productividad y reduce las emisiones de GEI.
La genética contra emisiones de CH4
Las emisiones de CH4 se pueden reducir teniendo una mejor eficiencia productiva a través de la selección genética: menor estrés calórico, mejor edad al destete o al sacrificio, menor incidencia de enfermedades; mejor selectividad de las plantas durante el pastoreo, mayor tasa de reproducción, tasa de pasaje del alimento en el rumen, entre otros (Clack et al., 2013; Knapp et al., 2014; Ross et al., 2013). Otra estrategia es la selección de rumiantes a través del consumo de alimento residual (RFI, h2 = 0.39) que tiene una heredabilidad media (Arthur et al., 2001). Dini et al. (2019) observaron una disminución en la emisión de metano del 26.8%. Se ha demostrado que herbivoros seleccionados por una tasa de pasaje rápida en el rumen y con alta cantidad de extracto libre de nitrógeno (ELN) disminuyen las emisiones de CH4 en un 21%, sin comprometer el rendimiento productivo (Okine et al., 2002). El progreso genético en los bovinos lecheros se ha conseguido en EU, al obtener una reducción en las emisiones de CH4 del 57% por unidad de producto de ECM (Capper et al., 2009). Se ha observado que las vacas jersey llegan a la madurez más rápido que las vacas Holstein, por lo tanto, la energía que requieren para el crecimiento es menor en la primera y segunda lactancia (Olson et al., 2010), esto indica que son más eficientes, con respecto a las vacas Holstein, durante las primeras lactancias. Las emisiones de CH4 son menores 8-12% en vacas de la raza Holstein comparadas con la raza New Zealand Friesian cuando consumen dietas mezcladas o pasturas (Robertson & Waghorn, 2002). La selección mediante razas especializadas puede aportar una mejora en el rendimiento productivo y en la emisión de CH4. Se puede alcanzar una reducción de las emisiones de CH4 para el 2050 (De Haas et al., 2021).
CONCLUSIONES
La disminución de las emisiones de CH4 en los sistemas pastoriles puede ser posible con la implementación de estrategias rigurosas en la nutrición y alimentación de rumiantes. Una depuración de unidades improductivas. Tener un plan de mejoramiento genético desarrollado con herbívoros adaptados a la región que toleren los desafíos durante el ciclo productivo que permita enfrentar el estrés calórico, sequias o enfermedades. La evaluación permanente de la capacidad de carga animal para realizar la rotación de potreros y el aprovechamiento de zacates nativos junto con plantas de valor forrajero como leguminosas y la inclusión de ensilados de cereales forrajeros. Por último, la adopción de una o más estrategias en el sistema productivo tendrá un mayor efecto en la mitigación de emisiones de CH4.
REFERENCIAS
Abecia, L., Toral, P. G., Martín-García, A. I., Martínez, G., Tomkins, N. W., Molina-Alcaide, E.,
Newbold, C. J., & Yáñez-Ruiz, D. R. (2012). Effect of bromochloromethane on methane
emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy
goats. Journal of Dairy Science, 95, 2027-2036. https://doi.org/10.3168/jds.2011-4831
Abreu, A., Carulla, J. E., Kreuzer, M., Lascano, C. E., Díaz, T. E., Cano, A., & Hess, H. D. (2003).
Efecto del fruto, del pericarpio y del extracto semipurificado de saponinas de Sapindus
saponaria sobre la fermentación ruminal y la metanogénesis in vitro en un sistema RUSITEC.
Revista Colombiana de Ciencias Pecuarias, 16, 147-154. https://www.redalyc.org/articulo. oa?id=295026126006
Anderson, R. C., Huwe, J. K., Smith, D. J., Stanton, T. B., Krueger, N. A., Callaway, T. R., Edrington,
T. S., Harvey, R. B., & Nisbet, D. J. (2010). Effect of nitroethane, dimethyl-2-nitroglutarate
and 2-nitro-methyl-propionate on ruminal methane production and hydrogen balance in vitro.
Bioresource Technology, 101, 5345-5349. https://doi.org/10.1016/j.biortech.2009.11.108
Appuhamy, J. A. D. R. N., Strathe, A. B., Jayasundara, S., Wagner-Riddle, C., Dijkstra, J., France,
J., & Kebreab, E. (2013). Anti-methanogenic effects of monensin in dairy and beef cattle: A
meta-analysis. Journal of Dairy Science, 96, 5161-5173. https://doi.org/10.3168/jds.2012-5923
Arthur, P. F., Renand, G., & Krauss, D. (2001). Genetic and phenotypic relationships among different
measures of growth and feed efficiency in young Charolais bulls. Livestock Production
Science, 68, 131-139. https://doi.org/10.1016/S0301-6226(00)00243-8
Baudracco, J., López-Villalobos, N., Holmesa, C. W., & Macdonald, K. A. (2010). Effects of stocking
rate, supplementation, genotype and their interactions on grazing dairy systems: a review. New
Zealand Journal o f Agricultural Research, 53, 109-133. https://doi.org/10.1080/00288231003777665
Benchaar, C., & Greathead, H. (2011). Essential oils and opportunities to mitigate enteric methane
emissions from ruminants. Animal Feed Science and Technology, 166, 338-355. https://doi.
org/10.1016/j.anifeedsci.2011.04.024
Brask, M., Lund, P., Weisbjerg, M. R., Hellwing, A. L. F., Poulsen, M., Larsen, M. K., & Hvelplund,
T. (2013). Methane production and digestion of different physical forms of rapeseed asfat
supplements in dairy cows. Journal of Dairy Science, 96, 2356-2365. https://doi.org/10.3168/
jds.2011-5239
Brown, E. G., Anderson, R. C., Carstens, G. E., Gutierrez-Bañuelos, H., McReynolds, J. L., Slay,
L. J., Callaway, T. R., & Nisbet, D. J. (2011). Effects of oral nitroethane administration on
enteric methane emissions and ruminal fermentation in cattle. Animal Feed Science and
Technology, 166(167), 275-281. https://doi.org/10.1016/j.anifeedsci.2011.04.017
COTECOCA. Comisión Técnico Consultiva para la determinación de Coeficientes de Agostadero (1967).
Metodología para determinar tipos vegetativos, sitios y productividad de sitios. Secretaría de
Agricultura y Recursos Hidráulicos Publicación No. 8, México, D.F. 84 p.
Capper, J. L., Cady, R. A., & Bauman, D. E. (2009). The environmental impact of dairy production: 1944
compared with 2007. Journal of Animal Science, 87, 2160-2167. https://doi.org/10.2527/jas.2009-1781
Carmona-Flores, L., Bionaz, M., Downing, T., Sahin, M., Cheng, L., & Ates, S. (2020). Milk
production, N partitioning, and methane emissions in dairy cows grazing mixed or spatially
separated simple and diverse pastures. Animals, 10(8), 1301. https://doi.org/10.3390/ani10081301
Clark, H. (2013). Nutritional and host effects on methanogenesis in the grazing ruminant. Animal,
7, 41-48. https://doi.org/10.1017/S1751731112001875
Crosson, P., Shalloo, L., O’Brien, D., Lanigan, G. J., Foley, P. A., Boland, T. M., & Kenny, D. A.
(2011). A review of whole farm systems models of greenhouse gas emissions from beef and
dairy cattle production systems. Animal Feed Science and Technology, 166-167, 29-45.
https://doi.org/10.1016/j.anifeedsci.2011.04.001
Chung Y. H., He, M. L., McGinn, S. M., McAllister, T. A., & Beauchemin, K. A. (2011). Linseed
suppresses enteric methane emissions from cattle fed barley silage, but not from those fed
grass hay. Animal Feed Science and Technology, 166-167, 321-329. https://doi.org/10.1016/j.
anifeedsci.2011.04.022
Chuntrakort, P., Otsuka, M., Hayashi, K., Takenaka, A., Udchachon, S., & Sommart, K. (2014).
The effect of dietary coconut kernels, whole cottonseeds and sunflower seeds on the intake,
digestibility and enteric methane emissions of Zebu beef cattle fed rice straw based diets.
Livestock Science, 161, 80-89. https://doi.org/10.1016/j.livsci.2014.01.003
DEFRA (Ministerio del Medio Ambiente, la Alimentación y los Asuntos Rurales del Reino Unido).
(2010). Ruminant nutrition regimes to reduce methane and nitrogen emissions. Project AC0209
Report. DEFRA. http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More
&Location=None&Completed=0&ProjectID=14952
Delgado, D. C., González, R., Galindo, J., Cairo, J., & Almeida, M. (2007). Potencialidad de
Trichantera gigantea y Morus alba para reducir la producción ruminal de metano in vitro. Revista
Cubana de Ciencia Agrícola, 41, 339-342. http://www.redalyc.org/articulo.oa?id=193017712007
De Haas, Y., Veerkamp, R. F., De Jong, G., & Aldridge, M. N. (2021). Selective breeding as a mitigation
tool for methane emissions from dairy cattle. Animal, 15, 100294. https://doi.org/10.1016/j.
animal.2021.100294
Díaz Puentes, L. N. (2009). Interacciones moleculares entre plantas y microorganismos: saponinas
como defensas químicas de las plantas y su tolerancia a los microorganismos. Una revisión.
RET. Revista de Estudios Transdisciplinarios, 1(2), 32-55. https://www.redalyc.org/pdf/1792/
Resumenes/Resumen_179214945004_1.pdf
Dini, Y., Cajarville, C., Gere, J. I., Fernandez, S., Fraga, M., Pravia, M. I., Navajas, E. A., & Ciganda,
V. S. (2019). Association between residual feed intake and enteric methane emissions in
Hereford steers. Translational Animal Science, 3(1), 239-246. https://doi.org/10.1093/tas/txy111
Foley, P. A., Kenny, D. A., Lovett, D. K., Callan, J. J., Boland, T. M., & O’Mara, F. P. (2009). Effect
of dl-malic acid supplementation on feed intake, methane emissions, and performance of
lactating dairy cows at pasture. Journal of Dairy Science, 92, 3258-3264. https://doi.
org/10.3168/jds.2008-1633
Galindo, J., Delgado, D. C., Pedraza, R., & García, D. E. (2005). Impacto de los árboles, los arbustos
y otras leguminosas en la ecología ruminal de animales que consumen dietas fibrosas. Pastos
y Forrajes, 28, 59-68. http://www.redalyc.org/articulo.oa?id=269121628005
Galindo, J., González, N., Delgado, D., Sosa, A., Marrero, Y., González, R., Aldana, A. I., & Moreira,
O. (2008). Efecto modulador de Leucaena leucocephala sobre la microbiota ruminal. Zootecnia
Tropical, 26, 249-252.
Galindo, J., González, N., Sosa, A., Ruiz, T., Torres, V., Aldana, A. I., Díaz, H., Moreira, O., Sarduy,
L., & Noda, A. C. (2011). Efecto de Tithonia diversifolia (Hemsl.) Gray (Botón de oro) en
la población de protozoos y metanógenos ruminales en condiciones in vitro. Revista Cubana
de Ciencia Agrícola, 45, 33-37. biblat.unam.mx/hevila/Revistacubanadecienciaagricola/
2011/vol45/no1/8.pdf
Garg, M. R. (2012). Balanced feeding for improving livestock productivity-increase in milk production
and nutrient use efficiency in methane emision (FAO Animal production and health paper
No. 173). FAO. http://www.fao.org/docrep/016/i3014e/i3014e00.htm
Gerber, P. J., Steinfeld, H., Henderson, B., Mottet, A., Opio, C., Dijkman, J., Falcucci, A., & Tempio,
G. (2013). Tackling climate change through livestock- A global assessment of emissions and
mitigation opportunities. FAO. http://www.fao.org/docrep/018/i3437e/i3437e00.htm
Goel, G., & Makkar, H. P. (2012). Methane mitigation from ruminants using tannins and saponins.
Tropical Animal Health and Production, 44, 729-739. https://doi.org/10.1007/s11250-011-9966-2
Grainger, C., & Beachemin, K. A. (2011). Can enteric methane emissions from rumiants be lowered
without lowering their production? Animal Feed Science and Technology, 166-167, 308-320.
https://doi.org/10.1016/j.anifeedsci.2011.04.021
Herrera-Pérez, J., Hernández-González, D., Sánchez-Santillán, P., Torres-Salado, N., Ayala-Monter,
M. A., Saavedra-Jiménez, L. A., & Rosales-Martínez, G. N. (2023). Características químicas
y fermentativas in vitro de pastos tropicales a diferentes edades de rebrote. Agrociencia,
57(6), 1102-1125. https://doi.org/10.47163/agrociencia.v57i6.2738
Hess, H. D., Monsalve, L. M., Lascano, C. E., Carulla, J. E., Diaz, T. E., & Kreuzer, M. (2003).
Supplementation of a tropical grass diet with forage legumes and Sapindus saponaria fruits:
effects on in vitro ruminal nitrogen turnover and methanogenesis. Crop and Pasture Science,
54, 703-713. https://doi.org/10.1071/AR02241
Holenchek, J. L., Pieper, R. D., & Herbel, C. H. (1989). Range management, principles and practices.
Prentice Hall.
Hristov, A. N., Oh, J., Lee, C., Meinen, R., Montes, F., Ott, T., Firkins, J., Rotz, A., Dell, C.,
Adesogan, A., Yang, W., Tricarico, J., Kebreab, E., Waghorn, G., Dijkstra, J., & Oosting, S.
(2013). Mitigation of greenhouse gas emissions in livestock production- A review of technical
options for Non-CO2 emissions (FAO Animal production and Health paper No. 177), FAO.
http://www.fao.org/docrep/018/i3288e/i3288e00.htm
Hristov, A. N., Oh, J., Giallongo, F., Frederick, T. W., Harper, M. T., Weeks, H. L., Branco, A. F.,
Moate, P. J., Deighton, M. H., Williams, S. R. O., Kindermann, M., & Duval, S. (2015). An
inhibitor persistently decreased enteric methane emissions from dairy cows with no negative
effect on milk production. Proceedings of the National Academy of Sciences, 112(34),
10663-10668. https://doi.org/10.1073/pnas.1504124112
IPCC. (2013). Climate change 2013: The physical basis. Contribution of working group I to the fifth
assessment report of the intergovernmental panel on climate, Geneva, Switzerland. http://
www.ipcc.ch/report/ar5/wg1/
Jahani-Azizabadi, H., Mesgaran, M. D., Vakili, A. R., Razayazdi, K., & Hashemi, M. (2011). Effect
of various medicinal plant essential oils obtained from semi-arid climate on rumen fermentation
characteristics of a high forage diet using in vitro batch culture. African Journal of. Microbiology
Research, 5(27), 4812-4819. https://doi.org/10.5897/AJMR11.575
Jouany, J. P., & Morgavi, D. P. (2007). Use of ‘natural’ products as alternatives to antibiotic feed additives
in ruminant production. Animal, 1, 1443-1466. https://doi.org/10.1017/S1751731107000742
Kim, D. H., Mizinga, K. M., Kube, J. C., Friesen, K. G., McLeod, K. R., & Harmon, D. L. (2014).
Influence of monensin and lauric acid distillate or palm oil on in vitro fermentation kinetics
and metabolites produced using forage and high concentrate substrates. Animal Feed Science
and Technology, 189, 19-29. https://doi.org/10.1016/j.anifeedsci.2013.12.010
Knapp, J. R., Laur, G. L., Vadas, P. A., Weiss, W. P., & Tricarico, J. M. (2014). Enteric methane in
dairy cattle production: Quantifying the opportunities and impact of reducing emissions.
Journal of Dairy Science, 97, 3231-3261. https://doi.org/10.3168/jds.2013-7234
Kobayashi, Y. (2010). Abatement of methane production from ruminants: trends in the maniplation
of rumen fermentation. Asian-Aust. Journal of Animal Science, 23, 410-416. https://doi.
org/10.5713/ajas.2010.r.01
Króliczewska, B., Pecka-Kiełb, E., & Bujok, J. (2023). Strategies used to reduce methane emissions
from ruminants: Controversies and issues. Agriculture, 13(3), 602. https://doi.org/10.3390/
agriculture13030602
Ku-Vera, J., Castelán-Ortega, O. A., Galindo-Maldonado, F. A., Arango, J., Chirinda, N., Jiménez-Ocampo,
R., Valencia-Salazar, S. S., Flores-Santiago, E. J., Montoya-Flores, M. D., Molina-Botero,
I. C., Piñeiro-Vázquez, A. T., Arceo-Castillo, J. I., Aguilar-Pérez, C. F., Ramírez-Avilés,
L., & Solorio-Sánchez, F. J. (2020). Review: strategies for enteric methane mitigation in
cattle fed tropical forages. Animal, 14(3), s453-s463. https://doi.org/10.1017/S1751731120001780
Lila, Z. A., Mohammed, N., Kanda, S., Kamada, T., & Itabashi, H. (2003). Effect of sarsaponin
on ruminal fermentation with particular reference to methane production in vitro. Journal of
Dairy Science, 86, 3330-3336. https://doi.org/10.3168/jds.S0022-0302(03)73935-6
Mao, H.-L., Wang, J. K., Zhou, Y. Y., & Liu, J. X. (2010). Effects of addition of tea saponins and
soybean oil on methane production, fermentation and microbial population in the rumen of
growing lambs. Livestock Science, 129, 56-62. https://doi.org/10.1016/j.livsci.2009.12.011
Min, B. R., Lee, S., Jung, H., Miller, D. N., & Chen, R. (2022). Enteric methane emissions and
animal performance in dairy and beef cattle production: Strategies, opportunities, and impact
of reducing emissions. Animals, 12(8), 948. https://doi.org/10.3390/ani12080948
Montenegro, J., & Abarca. S. (2000). Fijación de carbono, emisión de metano y de óxido nitroso en
sistemas de producción bovina en Costa Rica. En C. Pomareda & H. Steinfeld (Eds.),
Intensificación de la ganadería en Centroamérica: Beneficios económicos y ambientales.
CATIE- FAO-SIDE. Ed Nuestra Tierra. 334 p.
Moscoso, J. E., Franco, F., San Martín, F., Olazábal, J., Chino, L. B., & Pinares-Patiño, C. (2017). Producción de metano en vacunos al pastoreo suplementados con ensilado, concentrado y
taninos en el Altiplano Peruano en época seca. Revista de investigaciones veterinarias del
Perú, 28(4), 822-833. https://doi.org/10.15381/rivep.v28i4.13887
Murray, R. M., Bryant A. M., & Leng, R. A. (1976). Rates of production of methane in the rumen
and large intestines of sheep. British Journal of Nutrition, 36, 1-14. DOI:10.1079/BJN19760053
Mwenya, B., Santoso, B., Sar, C., Gamo, Y., Kobayashi, T., Arai, I., & Takahashi, J. (2004). Effects
of including β1–4 galacto-oligosaccharides, lactic acid bacteria or yeast culture on methan
genesis as well as energy and nitrogen metabolism in sheep. Animal Feed Science and
Technology, 115, 313-326. https://doi.org/10.1016/j.anifeedsci.2004.03.007
Nuñez, G. F. A. (2009). Fundamentos de crecimiento y evaluación animal. Trafford Publishing.
O’Mara, F. P. (2011). The significance of livestock as a contributor to global greenhouse gas emissions
today and in the near future. Animal Feed Science and Technology, 166-167, 7-15. https://
doi.org/10.1016/j.anifeedsci.2011.04.074
Olson, K. M., Cassell, B. G., & Hanigan, M. D. (2010). Energy balance in first-lactation Holstein,
Jersey, and reciprocal F 1 crossbred cows in a planned crossbreeding experiment. Journal of
Dairy Science, 93, 4374-4385. https://doi.org/10.3168/jds.2010-3195
Opio, C., Gerber, P., Mottet, A., Falcucci, A., Tempio, G., MacLeod, M., Vellinga, T., Henderson,
B., & Steinfeld, H. (2013). Greenhouse gas emissions from ruminant supply chains. A global
life cycle assessment. FAO.
Okine, E. K., Basarab, J. A., Baron, V., & Price, M. A. (2002). Methane and manure production in cattle with different net feed intake. Journal of Animal Science, 80(Suppl. 1), 206.
Patra, A. K. (2013). The effect of dietary fats on methane emissions, and its other effects on digestibility,
rumen fermentation and lactation performance in cattle: A meta-analysis. Livestock Science,
155, 244-254. https://doi.org/10.1016/j.livsci.2013.05.023
Pellikaan, W. F., Stringano, E., Leenaars, J., Bongers, D. J., van Laar-van, S., Schuppen, J., Plant,
& Mueller-Harvey, I. (2011). Evaluating effects of tannins on extent and rate of in vitro gas
and CH4 production using an automated pressure evaluation system (APES). Animal Feed
Science and Technology, 166, 377-390. https://doi.org/10.1016/j.anifeedsci.2011.04.072
Pinos-Rodríguez, J. M., García-López, J. C., Peña-Avelino, L. Y., Rendón-Huerta, J. A., González-González,
C., & Tristán-Patiño, F. (2012). Impactos y regulaciones ambientales del estiércol generado
por los sistemas ganaderos de algunos países de América. Agrociencia, 46, 359-370. http://
www.redalyc.org/articulo.oa?id=62990106
Pinares-Patiño, C. S., Waghorn, G. C., Hegarty, R. S., & Hoskin, S. O. (2009). Effects of intensification
of pastoral farming on greenhouse gas emissions in New Zealand. New Zealand Veterinary
Journal, 57, 252-261. https://doi.org/10.1080/00480169.2009.58618
Posada-Ochoa, S. L., Ramírez-Agudelo, J. F., & Rosero-Noguera, R. (2014). Producción de metano
y digestibilidad de mezclas kikuyo (Pennisetum clandestinum)-papa (Solanum tuberosum).
Agronomía Mesoamericana, 25, 141-150. https://doi.org/10.15517/am.v25i1.14214
Ramírez-Restrepo, C. A., & Barry, T. N. (2005). Alternative temperate forages containing secondary
compounds for improving sustainable productivity in grazing ruminants. Animal Feed
Science and Technology, 120, 179-201. https://doi.org/10.1016/j.anifeedsci.2005.01.015
Rivera, J. E., & Chará, J. (2021). CH4 and N2O emissions from cattle excreta: a review of main
drivers and mitigation strategies in grazing systems. Frontiers in Sustainable Food Systems,
5, 657936. https://doi.org/10.3389/fsufs.2021.657936
Robertson, L. J. & Waghorn, G. C. (2002). Dairy industry perspectives on methane emissions and
production from cattle fed pasture or total mixed rations in New Zealand. Proceedings of the
New Zealand Society of Animal Production, 62, 213-218 https://doi.org/10.3168/jdsc.2022-0297
Rojo-Martínez, G. E., Jasso-Mata, J., & Velásquez-Martínez, A. (2003). Las masas forestales como
sumideros de CO2 ante un cambio climático global. Revista Chapingo. Serie Ciencias Forestales
y del Ambiente, CIE. 9, 57-67. http://www.redalyc.org/articulo.oa?id=62990106
Ross, E. M., Moate, P. J., Marett, L., Cocks, B. G., & Hayes, B. J. (2013). Investigating the effect
of two methane-mitigating diets on the rumen microbiome using massively parallel sequencing
Journal of Dairy Science, 96, 6030-6046. https://doi.org/10.3168/jds.2013-6766
Steinfeld, H., Gerber, P., Wassenaar, T., Castel, V., Rosales, M., & de Haan. C. (2006). La larga sombra
del ganado. Problemas ambientales y opciones. FAO.
Steinfeld, H., & Gerber, P. (2010). Livestock production and the global environment: consume
less or produce better?. Proceedings of the National Academy of Science, 107(43),18237-18238.
https://doi.org/10.1073/pnas.1012541107
Thøy, K., Wenzel, H., Jensen, A. P., & Nielsen, P. (2009). Biogas from manure represents a huge
potential for reduction in global greenhouse gas emissions. IOP Conference Series: Earth
and Environmental Science, 6(24), 242020. https://doi.org/10.1088/1755-1307/6/4/242020
Tiemann, T. T., Lascano, C. E., Kreuzer, M., & Hess. H. D. (2008). The ruminal degradability of
fibre explains part of the low nutritional value and reduced methanogenesis in highly tanniniferous
tropical legumes. Journal of the Science of Food and Agriculture, 88, 1794-1803. https://doi.
org/10.1002/jsfa.3282
Van Gastelen, S., Antunes-Fernandes, E. C., Hettinga, K. A., Klop, G., Alferink, S. J. J., Hendriks,
W. H., & Dijkstra, J. (2015). Enteric methane production, rumen volatile fatty acid concentrations,
and milk fatty acid composition in lactating Holstein-Friesian cows fed grass silage-or corn
silage-based diets. Journal of Dairy Science, 98, 1915-1927. https://doi.org/10.3168/jds.2014-8552
Vigan, A., Lasseur, J., Benoit, M., Mouillot, F., Eugène, M., Mansard, L., Vigne, M., Lecomte, P.,
& Dutilly, C. (2017). Evaluating livestock mobility as a strategy for climate change mitigation:
Combining models to address the specificities of pastoral systems. Agriculture, Ecosystems
& Environment, 242, 89-101. https://doi.org/10.1016/j.agee.2017.03.020
Villa-Herrera, A., Paz-Pellat, F., Pérez-Hernández, M. J., Rojas-Montes, C., Rodríguez Arvizu, M.,
Ortiz-Acosta, S., Casiano-Dominguez, M., & Díaz-Solís, H. (2014). Estimación de la capacidad
de carga animal en agostaderos usando un índice de vegetación de pendientes normalizadas.
Agrociencia, 48, 599-614.
Villanueva, C., Ibrahim, M., & Haensel. (2010). Producción y rentabilidad de sistemas silvopastoriles:
Estudios de caso en América Central (Serie técnica-Manual técnico No. 95). Centro Agronómico
Tropical de Investigación y Enseñanza (CATIE).
Voet, D., Voet, J. G., & Pratt, C. W. (2007). Fundamentos de bioquímica (2a ed.) Médica Panamericana.
Waghorn, G., Tavendale, M., & Woodfield, D. (2002). Methanogenesis from forages fed to sheep.
Proceedings of the New Zealand Grassland Association, 64, 167-171. https://doi.org/10.33584/
jnzg.2002.64.2462
Wood, T. A., Wallace, R. J., Rowe, A., Price, J., Yáñez-Ruiz, D. R., Murray, P., & Newbold, C. J.
(2009). Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions.
Animal Feed Science and Technology, 152, 62-71. https://doi.org/10.1016/j.ics.2006.02.018